СОДЕРЖАНИЕ
|
стр. |
||
|
СПИСОК УСЛОВНЫХ СОКРАЩЕНИЙ |
4 |
|
|
ВСТУПЛЕНИЕ |
5 |
РАЗДЕЛ 1. ОБЗОР ЛИТЕРАТУРЫ
|
1.1. |
История исследования фенилкетонурии |
|
|
1.2. |
Картирование и клонирование гена фенилаланингидроксилазы |
|
|
1.3. |
Структура гена фенилаланингидроксилазы |
|
|
1.4. |
Белковый продукт гена ФАГ |
|
|
1.5. |
Полиморфные последовательности ДНК гена ФАГ |
|
|
1.6. |
Мутации гена фенилаланингидроксилазы |
|
|
1.7. |
Стадии ПЦР анализа |
|
1.8. |
Выдиление ДНК матрици |
|
1.9. |
Постановка ПЦР |
|
1.9.1. |
Компоненты реакционной смеси |
|
1.9.1.1. |
Праймеры |
|
1.9.1.2. |
Фермент. |
|
1.9.1.3. |
dNTP |
|
1.9.1.4. |
Буфер |
|
1.9.1.5. |
Концентрация Mg2+. |
|
1.9.1.6. |
Анализируемый образец |
|
1.9.1.7. |
ПЦР добавка |
|
1.9.2. |
Температурный режим |
|
1.9.3. |
Число циклов |
|
1.9.4. |
«Эффект плато» |
|
1.9.5. |
Способ постановки ПЦР |
|
1.9.5.1. |
Способ постановки ПЦР с использованием «горячего старта» |
|
1.9.5.2. |
«Nested» ПЦР |
|
1.9.5.3. |
Полуколичественный анализ |
1.9.6. |
Контроль за прохождением реакции амплификации |
|
1.9.6.1. |
Положительные контроли |
|
1.9.6.2. |
Внутренние контроли |
|
1.10. |
Детекция |
|
1.10.1. |
Метод горизонтального электрофореза |
|
1.10.2. |
Метод вертикального электрофореза |
|
1.11. |
Проблема контаминации |
РАЗДЕЛ 2. МАТЕРИАЛЫ И МЕТОДЫ исследований |
||
|
2.1. |
Материалы исследования |
|
|
2.2. |
Методы исследований |
|
|
2.2.1. |
Выделение и очистка ДНК |
|
|
2.2.1.1. |
Выделение и очистка лейкоцитарной ДНК из образцов свежей крови |
|
|
2.2.2.2. |
Выделение ДНК из образцов крови длительного хранения, а также некультивированных клеток амниотической жидкости, хориона и плаценты. |
|
|
2.2.2. |
Амплификация некоторых последовательностей гена ФАГ методом полимеразной цепной реакции |
|
|
2.2.3. |
Идентификация мутаций методом рестрикционного анализа |
|
|
2.2.3.1. |
Амплификация in vitro последовательностей гена ФАГ для последующего рестрикционного анализа |
|
|
2.2.3.2. |
Рестрикционный анализ |
|
|
2.2.4. |
Анализ полиморфных последовательностей ДНК гена ФАГ |
|
|
2.2.4.1. |
Анализ аллельных вариантов VNTR-локуса 3`-нетранслируемого участка гена ФАГ |
|
|
2.2.4.2. |
Анализ аллельных вариантов STR-локуса (3-й интрон) гена ФАГ |
|
|
РАЗДЕЛ 3. РЕЗУЛЬТАТЫ ИССЛЕДОВАНИй |
||
СПИСОК УСЛОВНЫХ СОКРАЩЕНИЙ
|
БСА |
- |
бычий сывороточный альбумин |
|
ДНК |
- |
дезоксирибонуклеиновая кислота |
|
ЭДТА |
- |
этилендиаминтетрааминуксусная кислота |
|
ПААГ |
- |
полиакриламидний гель |
|
ПДРФ |
- |
полиморфизм длины рестрикционных фрагментов |
|
ПЦР |
- |
полимеразная цепная реакция |
|
РНК |
- |
рибонуклеиновая кислота |
|
ФАГ |
- |
фенилаланингидроксилаза |
|
ФКУ |
- |
фенилкетонурия |
|
DGGE |
- |
денатурирующий градиентный гель-электрофорез |
|
DHPLC |
- |
высокоэффективная жидкостная хроматография |
|
dNTP, дНТФ |
- |
дезоксинуклеотидтрифосфат |
|
STR |
- |
короткие тандемные повторы (short tandeme repeat) |
|
VNTR |
- |
варьирующее число тандемных повторов (variable number tandem repeat). |
|
п.н. |
- |
пар нуклеотидов |
|
т. п. н. |
- |
тысяч пар нуклеотидов |
|
kD |
- |
килодальтон |
|
Tris |
трис-(оксиметил)-аминометан tris(hydroxymethyl)aminomethane C4H11O3N |
ВСТУПЛЕНИЕ
В связи с расшифровкой последовательности ДНК генома человека на передний план выходит вопрос выяснения функциональной роли генов. Наиболее перспективным подходом в изучении генов человека, в условиях, когда проведение экспериментов ограничено особенностями исследуемого объекта, является анализ генов обуславливающих наследственные заболевания. Уже к 2000 году на хромосомах человека картировано около 1000 генов, мутации которых приводят к различным наследственным заболеваниям [1]. Одним из первых в геноме человека был картирован и идентифицирован ген фенилаланингидроксилазы, мутации в котором обусловливают развитие заболевания с аутосомно-рецессивным типом наследования – фенилкетонурии [2]. Изучение молекулярно-генетической природы этого заболевания позволяет исследовать частоту, природу и механизмы распространения мутаций в геноме человека. Анализ ассоциации генотипа и клинических проявлений заболевания отражает фенотипический эффект генетических изменений, а также служит основой для актуальных и перспективных исследований функционирования и взаимодействия генов.
С практической точки зрения, исследования молекулярно-генетических основ наследственных болезней человека открывает новые возможности в диагностике, профилактике и лечении тяжелых патологий наследственной природы. Реальностью сегодняшнего дня является применение программ генетического тестирования и пренатальной ДНК-диагностики, многих заболеваний, а делом ближайшего будущего – внедрение в клиническую практику методов генотерапии.
Цель и задачи исследования. Цель работы – разработка технологий проведения ПЦР для амплификации in vitro последовательности ДНК седьмого эксона гена ФАГ.
Для достижения цели были поставлены основные задачи исследования:
1. Освоить базовые технологии ДНК анализа.
2. Создать банк образцов лейкоцитарной ДНК больных и их родственников из семей высокого риска фенилкетонурии из разных регионов Украины.
3. Разработать эфективные условия проведения амплификации последовательности ДНК седьмого эксона гена ФАГ.
Объектом исследования является ДНК человека полученая из лейкоцитов периферической крови, а предметом – методы анализа мутаций в седьмом эксоне гена ФАГ.
В работе использовались методы: выделения и очистки ДНК, рестрикционный анализ, полимеразная цепная реакция, гель-электрофорез.
Научная новизна полученных результатов.
Была разработана матиматическая модель отжига праймеров которая позволила выбрать оптимальный температурный режым амплификации.
Практическое значение полученных результатов. Была разработана и оптимизирована методика для диагностики мутаций локализованых в последовательности седьмого экзона гена ФКУ.
Разработана стратегия использования прямого анализа мутаций гена ФАГ, а также анализа сцепления мутантного гена с высокоинформативными ДНК-полиморфными маркерами для пренатальной диагностики, массового и селективного скрининга гетерозиготных носителей мутантных генов с целью профилактики этого тяжелого наследственного заболевания в Украине.
Автор высказывает искреннюю благодарность сотрудникам отдела генетики человека ИМБИГ НАНУ за постоянную поддержку и помощь во время выполнения работы.
Объем и структура работы. Диплом состоит из вступления, обзора литературы, описания материалов и методов исследований, полученных результатов собственных исследований и их обсуждения, заключения, выводов и перечня использованных источников. Текст диплома проиллюстрирован таблицами и рисунками. Дипломная работа изложена на страницах машинописного текста.
Раздел 1
1.1 . История исследования природы фенилкетонурии
Фенилкетонурия – одно из наиболее распространенных наследственных заболеваний группы энзимопатий моногенной природы с аутосомно-рецессивным типом наследования.
Впервые в медицинской литературе описание этого заболевания появилось в 1934 году. [3]
Клинические симптомы фенилкетонурии появляются уже в первые месяцы жизни и могут быть разделены на такие группы: интеллектуальный дефект – умственная отсталость (чаще в степени имбицильности или глубокой дебильности); нейродегенеративные проявления; судорожный синдром [4].
Среди других симптомов выделяют: сходящееся и расходящееся косоглазие, асимметрия лица, снижение глоточного и нёбного рефлексов; вегетативно-трофические расстройства: сухость кожи, нарушение ее пигментации, гипергидроз дистальных отделов конечностей, ломкость волос, тенденция к артериальной гипотонии [5].
Основным диагностическим признаком заболевания является повышенная концентрация фенилаланина в крови, которая определялась ранее методом Гатри, а теперь методом иммунохимического анализа. В норме у детей этот показатель составляет 62±18 μM/л, а у больных ФКУ 120- 2400 μM/л [1]. В большинстве стран мира разработаны программы скрининга новорожденных на фенилкетонурию. В Украине эта программа действует, начиная с 1971. Благодаря этим программам не только стало возможным начинать пресимптоматическое лечение больных, но и определить частоту этого заболевания в разных странах. Так максимальная частота зарегистрирована в Ирландии 1:4500, а минимальная в Японии 1:120000 [6, 7]. В Украине на 8300 новорожденных рождается один ребенок больной ФКУ [Нечипоренко М. В. Исследование мутаций и полиморфизмов последовательности ДНК гена фенилаланингидроксилазы в семьях высокого риска фенилкетонурии. Диссертация на соискание ученой степени кандидата биологических наук], причем в разных регионах частота ФКУ существенно отличается. Например в Криворожской области 1:12000, а в Полтавской и Черниговской областях 1:4419. Частота носителей гена ФКУ в Украинской популяции 1:43 [9].
L-фенилаланин принадлежит к числу незаменимых аминокислот. В нормальных условиях лишь незначительная его часть используется для построения собственных белков, а большая часть превращается в тирозин [4].
В 1953 году Джервис и соавторы показали, что у больных фенилкетонурией преобразование фенилаланина в тирозин почти не происходит [10]. Позже было установлено, что причиной этого является отсутствие или значительное снижение активности лабильной (печеночной) фракции фермента фенилаланингидроксилазы [11]. Вследствие этого концентрация фенилаланина во всех жидких средах организма возрастает в десятки раз. В противоположность фенилаланину концентрация тирозина снижается [12].
Реакция превращения фенилаланина в тирозин осуществляется специальной ферментативной системой - гидроксилазы L-фенилаланина (ФАГ, фенилаланин 4-монооксигеназа, ЕС 1.14.16.1), которая катализирует присоединение гидроксильной группы к фенилаланину. В состав фермента входят два компонента: лабильный и стабильный [13, 14]. Стабильная фракция синтезируется во многих органах, в том числе почках, сердце, печени; лабильная фракция синтезируется только в печени [5]. В реакции преобразования фенилаланина в тирозин принимают участие кислород и тетрагидробиоптерин, который выступает в роли кофактора, а также дигидроптеридинредуктаза (ДГПР, КФ 1.6.99. 7) [13, 14, 15, 16, 17]. Реакция происходит по следующей схеме:
фенилаланин + О2 + тетрагидробиоптерин![]() тирозин + вода +
тирозин + вода +
окисленный биоптерин [18].
Эта реакция происходит при участии лабильной фракции фенилаланингидроксилазы. Стабильная часть ферментативной системы, а также ДГПР выполняют функцию восстановителя окисленного биоптерина в тетрагидробиоптерин [14].
У больных фенилкетонурией значительная часть фенилаланина выводится из организма с мочой, кроме того, для снижения концентрации фенилаланина происходит активация побочного пути преобразования аминокислоты – переаминирование. Около 10 % фенилаланина, поступающего с пищей, у больных фенилкетонурией выделяется с мочой в виде чистого фенилаланина, 30 % - в виде фенилпировиноградной кислоты (ФПК), 20 % - фенилмолочной кислоты и 10 % - фенилуксусной [19].
Первичный дефект синтеза фенилаланингидроксилазы приводит к накоплению в организме человека фенилаланина и его производных. Следствием этого является ингибирование ряда ферментов, что влечет за собой вторичные нарушения обмена важных аминокислот, в первую очередь тирозина и триптофана [4].
Тирозин принимает участие в строительстве чрезвычайно важных белков организма, а именно меланина, норадреналина, адреналина, тироксина, дофамина [20, 21]. На первый взгляд видно, что дефицита тирозина, и как следствие веществ, которые из него получаются, при ФКУ не должно быть, поскольку тирозин поступает в организм с пищей. Однако фенилаланин и его аномальные метаболиты ингибируют ферменты, которые принимают участие в обмене тирозина и триптофана [22].
1.2. Картирование и клонирование гена фенилаланингидроксилазы
Работы по картированию гена ФАГ были начаты еще в 70-е годы. Первоначально, картирование проводилось на основе анализа сцепления с другими полиморфными белковыми маркерами человека. Было показано, что ген ФАГ сцеплен с геном PGM-1 (фосфоглюкомутаза), а также генами AMY-1 AMY-2 (амилазные локусы) картированными на хромосоме 1 [23, 24]. Однако, в 1979 году метод анализа сцепления был усовершенствован и повторно проведенные исследования сцепления с другими маркерами не подтвердили эту информации [25].
Точно локализовать ген ФАГ удалось лишь в 1984 году, когда стало возможно применять технологии рекомбинатных ДНК. Лидски и соавторы гибридизировали по Саузерну ДНК-панели клеточных гибридов человек/мышь с полученными к тому времени клонами кДНК гена ФАГ человека. Максимальный гибридизационный сигнал был получен для клонотеки хромосомы 12. В результате последующей гибридизации in situ с хромосомой 12, у которой были делетированы различные участки, ген ФАГ был точно картирован в области 12q22-24.1 [2].
В 1982 году Робсон методом иммунопреципитации полисом выделил мРНК гена ФАГ печени кролика и клонировал кДНК [26]. В 1985 году Ву и соавторы, используя клонированную кДНК кролика в качестве специфической гибридизационной пробы, выделили и клонировали кДНК печени человека (клон phPAH47), на следующий год тем же коллективом авторов была определена полная последовательность нуклеотидов кДНК, а еще через год и полная последовательность гена ФАГ [27, 28].
Библиотека кДНК клеток печени человека, содержащая 14 миллионов рекомбинантов была сконструирована в клонируемом экспрессирующемся векторе lgt11. Для получения полной последовательности кДНК гена ФАГ авторами было проскринированo 100 000 рекомбинантов и получено 100 положительных гибридизационных сигналов. Четыре клона имели размер встроенного фрагмента 2000 - 2500 п.н., соответствующий полученному ранее методом гибридизации по Нозерну размеру мРНК фенилаланингидроксилазы. Эти 4 клона были обработаны рестриктазой EcoR1, встроенный фрагмент был извлечен и встроен по EcoR1 сайту плазмиды pBR322. Используя метод сиквенса, предложенный Максамом и Гилбертом, авторы определили размер и последовательность нуклетидов кДНК гена ФАГ человека [27, 28]
1.3. Структура гена фенилаланингидроксилазы
Ген фенилаланингидроксилазы имеет размер 90 т.п.н. В состав гена входит 13 экзонов и интронов [27]. Средняя длина экзонов приблизительно 114 п.н., самый короткий 9-ый экзон - 57 п.н., наиболее длинный 6-ой - 197 п.н. Тринадцатый экзон состоит из 894-х нуклеотидов, но протеинкодирующий участок содержит лишь 42 нуклеотида. Размеры интронов значительно больше: минимальная длина интрона 1т.п.н.(седьмой интрон), максимальная - 23.5 т.п.н. (третий интрон). В 3'-части гена сосредоточены более компактные интроны, в 5'-части - интроны больших размеров. Таким образом, суммарная длина экзонных последовательностей составляет 2.3 т.п.н., а интронных 85 т.п.н. Показатель соотношения некодирующей части гена к кодирующей для гена ФАГ один из наибольших в сравнении с другими известными на сегодня генами эукариот [27].
Было показано, что нуклеотидная последовательность гена фенилаланингидроксилазы человека имеют 90%-ю гомологию с аналогичным геном кролика [29] и 92,3% гомологию с геном ФАГ мыши [30], более того некоторые из описанных на сегодняшний день мутаций гена ФАГ человека идентифицированы и у мыши [31].
Экспрессия гена ФАГ происходит только в клетках печени. Незначительный уровень экспрессии отмечают уже в процессе эмбриогенеза, начиная с первого триместра беременности, но система гидроксилирования ФА включается в работу только в постнатальный период [32].
1.4. Белковый продукт гена ФАГ
Белковым продуктом гена ФАГ является железосодержащий фермент - фенилаланингидроксилаза, состоящий из 452-х аминокислотных остатков. Модель структуры фермента была создана в 1998-1999 г.г (рисунок 1.1) [33, 34]. Было показано, что фермент функционирует в виде гомотетрамера, молекулярная масса каждой субъединицы которого составляет 50-53 кD [35, 36]. Субъединицы фермента состоят из трех доменов: регуляторный домен (аминокислоты 1-142), каталитический домен (аминокислоты 143-406) и тетрамеризующий домен (аминокислоты 407-452) [32, 33, 34].
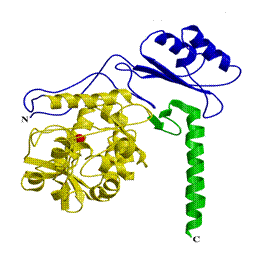
Рис. 1.1. Структура белка фенилаланингидроксилазы человека (модель). Красный - лиганд железа, желтый - католитический домен, фиолетовый - регуляторный домен, зелёный – тетромеризующий домен [33, 34, 35].
1.5. Полиморфные последовательности ДНК гена ФАГ
При изучении молекулярной структуры гена ФАГ было идентифицировано несколько полиморфных участков ДНК, которые можно разделить на следующие группы: 1) биаллельные полиморфизмы длины рестрикционных фрагментов (ПДРФ); 2) мультиаллельные полиморфизмы, представленные в последовательности гена ФАГ минисателлитным VNTR (варьирующее число тандемных повторов) и микросателлитным STR (короткие тандемные повторы) полиморфизмами; 3) однонуклеотидные полиморфизмы (SNP).
Исторически первыми были идентифицированы ПДРФ полиморфизмы гена ФАГ. В процессе исследования кДНК было найдено восемь полиморфных сайтов рестрикции для семи разных эндонуклеаз: BglI, PvuII, EcoRI, XmnI, MspI, EcoRV, HindIII [38]. На рисунке 1.2 представлен весь спектр и локализация полиморфизмов гена ФАГ. Точная локализация этих участков была проведена с помощью метода блот-гибридизации с кДНК-зондами, которые представляют собой разные участки гена.
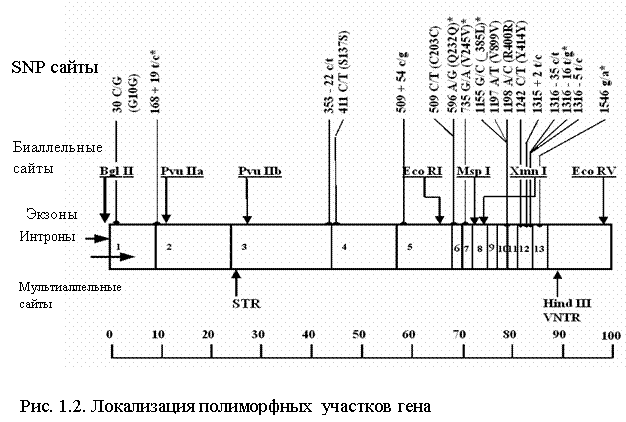
Полиморфизмы длины рестрикционных фрагментов (ПДРФ) ФАГ локуса образуют два кластера на 5'-конце и 3'-конце гена, расстояние между которыми приблизительно 30 т.п.н. В состав 5'-кластера входят экзоны 1-3 с полиморфными сайтами для эндонуклеаз BglI , PvuII (а), PvuII(б). Его размер составляет приблизительно 40 т.п.н. 3'-кластер с полиморфными сайтами для эндонуклеаз EcoRI , XmnI, MspI, EcoRV, HindIII включает экзоны 6-13 и имеет размер 30 т.п.н.[27]
При составлении рестрикционной карты гена ФАГ для HindIII полиморфизма, локализованного в 3`-части гена, было отмечено существование трех аллельных вариантов – явление не типичное для биаллельных ПДРФ полиморфизмов [38, 39]. Были определены такие размеры рестрикционных фрагментов, как HindIII 3,4 т.п.н.; HindIII 3,6 т.п.н. и HindIII 3,8 т.п.н. [40]. Для дальнейшего изучения этого участка гена ФАГ ДНК-зонды, содержащие 13-й экзон и 3 т.п.н. 3`-нетранслируемой последовательности проклонировали в ранее сконструированной плазмиде pKSP23E12. Это позволило более точно локализовать полиморфный регион и ограничить его участком размером 0,6 т.п.н [41]. При секвенировании этого фрагмента определили его первичную структуру и показали наличие АТ-богатого (70%) минисателлита на расстоянии 3 т.п.н от 13-го экзона в 3`-нетранслируемом участке. Размер повторяющейся единицы этого VNTR-полиморфизма составляет 30 п.н. Количество тандемных повторов варьирует от 3-х до 12-ти. Таким образом, полученные результаты объясняют наличие трех аллелей HindIII полиморфизма существованием минисателлитного VNTR полиморфизма в этом регионе, аллель с тремя повторами которого соответствует HindIII 3,4; 5; 6, 7, 8, 9 повторов – HindIII 3,6 и 12 повторов – HindIII 3,8 [41].
Для большинства мутаций гена ФАГ была показана ассоциация с определенными аллелями VNTR-полиморфизма, что свидетельствует о том что, какие бы события не привели к возникновению мультиаллельности в этом локусе они предшествовали процессу возникновения известных на сегодняшний день мутаций [41].
В транскрибируемой последовательности гена ФАГ идентифицировано 40 SNP полиморфизмов. Эти полиморфизмы представляют собой единичные нуклеотидные замены, но не являются ПДРФ. Девять SNP локализованы в интронных последовательностях и 31 - в экзонных [37]. Эти полиморфизмы называют молчащими, так как замена одного нуклеотида в аминокислоте при SNP полиморфизме не приводит к возникновению мутации, например Q232Q.
1.6. Мутации гена фенилаланингидроксилазы.
Как отмечалось выше, причиной развития ФКУ является дефект синтеза фермента ФАГ, а это, в свою очередь, является следствием мутаций в гене ФАГ. К 2001 году были идентифицированы более 600 мутаций, подавляющее большинство которых относится к классу миссенс мутаций, то есть мутации, которые приводят к единичной аминокислотной замене. Кроме миссенс мутаций есть нонсенс мутации, которые приводят к возникновению стоп кодона, а также делеции и мутации донорных и акцепторных сайтов сплайсинга [37]. На рисунке 1.3 представлена диаграмма, иллюстрирующая спектр мутаций каждого типа, известных к 2000 году [37].
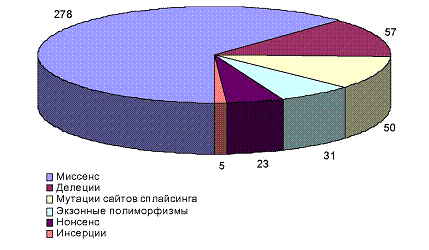
Рис. 1.3. Спектр мутаций разных типов, идентифицированных в гене ФАГ.
К 2001 году было идентифицировано 140-ка мутаций гена ФАГ. Распростронение мутаций по месту локализации представлена в татлице 1.1 [37]. Так как наибольшее число мутаций находится в седьмом экзоне, то седьмой экзон требует особого внимания. Наш интерес к седьмому экзону усиливается тем фактом, что при помощи одной амплификации и 6 рестрикций мы можем индифицировать 6 мутаций. Также представляет интерес такие распространеные мутации как: R408W, R261Q, G272X, P281L, R261X, S273F. Существует много методов анализа этих мутаций такие как: ПЦР, ИФА и блот гибридизация. Так как наименьшая сибестоимость одного анализа, чуствительность и точность достигается при помощи полимеразной цепной реакции, то мы отдадим предпочтение именно ПЦР.
Татлица 1.1. Распростронение мутаций по месту локализации.
|
Локализация |
Количество мутаций |
|
1 экзон |
7 |
|
3 экзон |
38 |
|
5-м экзоне |
1 |
|
7-м экзоне |
69 |
|
11-м экзон |
39 |
|
12 экзон |
22 |
|
10-м интрон |
1 |
|
12-м интрон |
1 |
1.7. ПЦР-анализ
ПЦР-анализ состоит из следующих этапов:
- выдиление ДНК матрици;
- амплификация;
- детекция амплифицированых продуктов;
- полный или частичный анализ нуклеотидной последовательности продуктов ПЦР такими методами как: рестрикционный анализ, гетеродуплексный анализ, электрофоретическое фракционирование.
1.8. Выдиление ДНК матрици
При подготовке пробы к постановке ПЦР используют различные методики в зависимости от поставленных задач. Их суть заключается в экстракции (извлечении) ДНК из биопрепарата и удалении или нейтрализации посторонних примесей для получения препарата ДНК с чистотой, пригодной для постановки реакции амплификации.
Иногда бывает достаточно прокипятить образец в течение 5-10 минут, однако в большинстве случаев требуются более трудоемкие методы.
Стандартной и ставшей уже классической считается методика получения чистого препарата ДНК, предложенная Marmur. Она включает в себя ферментативный протеолиз клеток с последующей депротеинизацией и переосаждением ДНК спиртом. Однако это метод довольно трудоемок и предполагает работу с такими агрессивными и имеющими резкий запах веществами, как фенол и хлороформ.
Одним из популярных в настоящее время является метод выделения ДНК, предложенный Boom с соавторами. Этот метод основан на использовании для лизиса клеток сильного хаотропного агента - гуанидина тиоционата (GuSCN) и последующей сорбции ДНК на носителе (стеклянные бусы, диатомовая земля, стеклянное «молоко» и т.д.). После процесса очистки в пробе остается ДНК, сорбированная на носителе, с которого она легко снимается с помощью элюирующего буфера. Метод удобен, технологичен и пригоден для подготовки образца к амплификации. Однако возможны потери ДНК вследствие необратимой сорбции на носителе, а также в процессе многочисленных очисток. Особенно большое значение это имеет при работе с небольшими количествами ДНК в образце. Кроме того, даже следовые количества GuSCN могут ингибировать ПЦР. Поэтому при использовании этого метода очень важен правильный выбор сорбента и тщательное соблюдение технологических нюансов. Следует отметить, что из-за большого количества стадий добавления и удаления растворов при работе с образцом требуется аккуратность, т.к. возможна перекрестная контаминация между пробами образующейся аэрозолью ДНК.
Другая группа методов пробоподготовки основана на использовании ионообменников типа Chilex, которые, в отличие от стекла, сорбируют не ДНК, а, наоборот, примеси, мешающие реакции. Как правило, эта технология включает две стадии: кипячение образца, в результате чего клеточные стенки разрушаются, а нуклеиновые кислоты выходят в раствор; и сорбция примесей на ионообменнике. Метод чрезвычайно привлекателен простотой исполнения. В большинстве случаев он пригоден для работы с клиническим материалом. К сожалению, иногда встречаются образцы с такими примесями, которые невозможно удалить с помощью ионообменников. Кроме того, клеточные стенки некоторых микроорганизмов не поддаются разрушению простым кипячением. В этих случаях необходимо введение дополнительных стадий обработки образца.
При массовом скрининге, когда важно получить статистические данные, допускается использование простых методов с применением детергентов или обработки биологического материала щелочами с последующей их нейтрализацией. В то же время, использование подобных методов пробоподготовки для клинической диагностики может приводить к ложноотрицательным результатам вследствие использования в реакционной смеси некачественного препарата ДНК.
Таким образом, к выбору метода пробоподготовки следует относиться с пониманием целей проведения предполагаемых анализов.
1.9. Постановка ПЦР
Полимеразная цепная реакция – это метод, имитирующий естественную репликацию ДНК и позволяющий обнаружить единственную специфическую молекулу ДНК в присутствии миллионов других молекул.
Открытие метода полимеразной цепной реакции (ПЦР) стало одним из наиболее выдающихся событий в области молекулярной биологии за последние десятилетия. Это позволило поднять медицинскую диагностику на качественно новый уровень.
Основные принципы использования праймеров (коротких искусственно синтезированных молекул ДНК) и состав ингредиентов, входящих в реакционную смесь для получения копий ДНК впервые были описаны Kleppe с соавт. в 1971 году. Однако тогда еще не была продемонстрирована основная черта ПЦР - экспоненциальное увеличение количества копий фрагмента исходной ДНК.
В 1983 году сотрудник фирмы «Cetus» Kary Mullis предложил метод, ставший в дальнейшем известным как полимеразная цепная реакция. Суть метода заключается в многократном копировании (амплификации) в пробирке определенных участков ДНК в процессе повторяющихся температурных циклов. На каждом цикле амплификации синтезированные ранее фрагменты вновь копируются ДНК-полимеразой. Благодаря этому происходит многократное увеличение количества специфических фрагментов ДНК, что значительно упрощает дальнейший анализ.
Следует отметить, что этому открытию сопутствовало развитие некоторых технологий. В частности, появление приборов, позволяющих автоматически синтезировать одноцепочечные фрагменты ДНК (олигонуклеотиды). В тот же период были обнаружены уникальные микроорганизмы, живущие в гейзерах. Их ферментативная система, в частности ДНК-полимераза, выдерживает высокие температуры горячих источников и сохраняет свою биологическую активность вплоть до 95°С, что является необходимым условием для проведения полимеразной цепной реакции.
Результатом открытия ПЦР стало почти немедленное практическое применение метода. В 1985 году Saiki с соавт. опубликовали статью, в которой была описана амплификация геномной последовательности ß-глобина. С этого момента количество публикаций, в которых авторы сообщали о применении ПЦР в своих работах, стало увеличиваться в геометрической прогрессии. Метод приобрел такую популярность, что сегодня уже трудно представить работу в области молекулярной биологии без его использования. Особенно бурное развитие метод полимеразной цепной реакции получил благодаря международной программе «Геном человека». Были созданы современные лазерные технологии сиквенирования (расшифровки нуклеотидных последовательностей ДНК). Если в недавнем прошлом для расшифровки последовательности ДНК размером в 250 пар нуклеотидов (п.н.) требовалась неделя, то современные лазерные сиквенаторы позволяют определять до 5000 п.н. в день. Это в свою очередь способствует значительному росту информационных баз данных, содержащих последовательности ДНК различных биологических объектов. В настоящее время предложены различные модификации ПЦР, показана возможность создания тест-систем для обнаружения микроорганизмов, выявления точечных мутаций, описаны десятки различных применений метода.
